During an educational research experience between 1993 and 1997, I came across several conceptual problems in applied ecology which eventually led me to confront the highly theoretical question Schrodinger (1944) posed nearly six decades ago - "What is life?" I offer the story here, as well as my current working hypothesis on the original and fundamental nature of life, as I believe both to be relevant to theory and application of ecological science.
While working towards a master's degree, I struggled to develop an indicator of ecological health for soils. I sought a scientifically-based, quantitative measure of soil biota by which to place a given agricultural soil on a continuum of "good" to "bad" health. This hybrid qualitative-quantitative metric of soil health was to be defined relative to the human societal values of 1) production of food and fiber and 2) sustainability of this agricultural production. Essentially, these societal values answered the question "good or bad (soil health) for what?" (Hess et al. 2000). In order to bridge the gap between objective, quantitative measurement and a subjective, qualitative value assessment, I sought a set of first principles capable of providing benchmark or reference values for what is important, what is good versus bad for production and sustainability from an ecological perspective. I worked with free-living (i.e., not plant parasitic) nematode communities due to their abundance, functional diversity and significant roles in decomposition and nutrient cycling processes.
The first approach I tried was to identify a suitable natural system - a reference site - by which to set a standard for good ecological health. This followed the work of Karr and his index of biotic integrity (IBI, Karr 1991). While I saw how this worked for streams and their biota, the reference site approach was problematic for agricultural systems because of the major differences in disturbance, inputs, and biodiversity between natural and highly managed systems. I saw a related problem in reports of manipulation experiments in which soils were completely sterilized and then biota allowed to recolonize (Ettema and Bongers 1993). Within a year or two, succession led to diverse and productive soil food webs. This amazing resilience of soil systems threatened an infinite regress in time when defining ecological health - that no matter how severely they are disturbed, soils could always return to a diverse and productive state. The issue of recolonization from surrounding undisturbed areas suggested an infinite regress in space - no matter how big an area of disturbance, there would always be some refuge from which biota could mount a comeback. I encountered a third seeming boundlessness in the concepts and topics of ecology itself - how to draw the line at nematode communities and their functions, to the exclusion of other soil fauna and flora, physical and chemical dynamics, etc., etc. ad infinitum. The combination of seeming limitlessness in space, time and ecological dynamics lead me to search for conceptual closure within which to locate a metric of ecological health.
In order to graduate, I postponed addressing what I sensed to be a very fundamental issue. Rather than develop an absolute reference basis for health, I ranked the relative sensitivity of nematode genera to physical and chemical aspects of agricultural management. I then used these sensitivity rankings to measure disturbance to the nematode community (Fiscus and Neher, in press). The interesting result from this approach was that it showed how effects of tillage, fertilizer and pesticides on nematode communities are a mixed bag - some genera and functional groups suffer while others are stimulated. But the encounter I had with the dilemmas of ecological health started a curiosity that lasted beyond my master's degree work. Reading broadly in pursuit of a conceptual bedrock basis for health, I eventually hit the question, "What is Life?" Since confronting this question (and driven to close up those infinite regresses in space, time and ecological interactions to make ecological health a meaningful concept) I have arrived at a place I did not expect - I have pieced together a suggestion of a fundamentally different understanding of life than that with which I began. From this alternative set of assumptions, I now see monitoring and assessment of ecological health in a different light.
The purpose of this paper is to present briefly the ecosystemic life hypothesis as compared and contrasted with the predominant organismic life approach. This hypothesis states that the ecosystemic organization - an autotroph functionally integrated with a heterotroph - is the original, general and self-perpetuating unit of life. By comparison, cells and organisms are seen as special case subunits of life that cannot persist in isolation. I suggest that the ecosystemic conceptualization of life fixes the problems of infinite boundlessness in space, time and interactions when attempting to assess ecological health - it offers strong (but still incomplete) closure and solid first principles for a standard of values of living systems. When one assumes that the fundamental and distinguishing characteristic of life is the capacity for sustained production - a merger of the two values which my indicator of soil health was to address - then an ecosystemic definition of life is preferable. I see sustained production as the key to open-ended evolution and simple core functions of an original ecosystem - molecular string composition and decomposition - as more general and more robust than cellular functions for understanding living systems and the origin and evolution of life. Thus, the iteration of simpler ecosystemic processes may have preceded and generated much more complex cells and organisms. I present this hypothesis as an amalgam of ideas provided by Lotka, Odum, Ulanowicz, Rosen and others.
A preliminary task will help to start - to describe existing definitions for life and for ecosystem. Solomonoff (1997) suggests the importance of operational definitions for science:
"An operational definition of anything is a precise sequence of physical operations that enable one to either construct it or identify it with certainty. When one can't make an operational definition of something, this is usually an indication of poor understanding of it. Attempts to operationalize definitions can be guides to discovery. I've found this idea to be an invaluable tool in telling me whether or not I really understand something."
I admit my attempts here do not produce fully operational definitions of life or ecosystem, but I suggest that the exercise itself is crucial for understanding both, is important for life science and helps define ecological health.
I think the basic dictionary definitions of life and ecosystem are the ones most often assumed and used. The Merriam-Webster Collegiate dictionary (online reference) defines life as "an organismic state characterized by capacity for metabolism, growth, reaction to stimuli, and reproduction." The same source defines ecosystem as "the complex of a community of organisms and its environment functioning as an ecological unit." Morowitz (1992) states that "all life is cellular in nature... A cell is the most elementary unit that can sustain life" however, see below). It seems fair to say that organisms are generally seen as fundamental (and indivisible) units of life, and ecosystems as interacting and higher order (and divisible) collections of these units.
In practice, the definitions are more complex and the distinctions are often blurred. The Encyclopedia Britannica (online reference) lists five different approaches to the definition of life based on physiological, metabolic, biochemical, genetic and thermodynamic considerations. O'Neill et al. (1987) speak to the fuzzy boundaries between life and ecosystem definitions; they "...define ecosystems as the smallest units that can sustain life in isolation from all but atmospheric surroundings. However, one is still left with the problem of specifying the area that should be included." This is akin to defining the ecosystem as the fundamental unit of life, as "smallest units" can be considered the simplest or most basic units. Morowitz (1992) also blends the two concepts, stating that "sustained life is a property of an ecological system rather than a single organism or species."
One potential avenue for clarification is to move to the framework of evolution, arguably the strongest basis for integration of biology and ecology. Eigen (1995), for example, tackled the definition of life by defining a unit of selection. He lists self-reproduction, mutation and metabolism as properties of systems that are "predestined" to selection, as in possessing the capacity for selection inherently. While these properties more often are associated with cells than ecosystems, his approach leaves open another question. Could an ecosystemic organization of physical and chemical dynamics provide a more robust unit of selection than a cellular, organismic, metabolic or genetic organization? This is another way to state the question to which ecosystemic life is a response. I believe the ecosystemic paradigm for the identification of life comes closer to an operational definition as it is more amenable to explanation in physical/chemical and biological dynamics simultaneously.
An interesting brief picture of the evolution of the ecosystem concept is provided by Allen and Roberts (in the foreword to Ulanowicz 1997). They chart the change in the concept as developed by Transeau and Tansley through Lindeman, E.P. Odum and Ulanowicz. Their story begins with the assertion of Tansley that a systems view of life treat environmental fluxes and biological fluxes together as a "reciprocity of influence". Lindeman and E.P. Odum then deepen the role of energy flux, and Ulanowicz distilled this yet again to focus on the non-conserved units of thermodynamics (entropy and exergy) as the key dynamic flux of ecosystems. A reciprocal relation with environment is arguably as important for understanding life as it is for understanding ecosystems. This, combined with the historical scientific trend to explain ecosystems in ever more physical terms, says to me that the concepts of life and ecosystem are necessarily linked and naturally convergent.
In the next article in this series, I present four concepts that I have found helpful in resolving the dilemmas of ecological health. Taken together, works by Lotka, H.T. Odum, Ulanowicz and Rosen also suggest a path toward a revised set of first principles for ecology.
In the first article in this series (Fiscus 2001), I introduced the ecosystemic life hypothesis as a potential solution to conceptual roadblocks I encountered when trying to define ecological health. I also showed how definitions of life and ecosystem are often blurred, and suggested that this lack of clear operational definitions for fundamental concepts hints at the convergence of ecosystem science with big questions such as "What is Life?" In this second article, I present four concepts that weave life and ecosystem together further and explain why they may be inextricably linked. Taken together or "connected", these concepts suggest a path toward a revised set of first principles for ecology. I present them separately and then attempt to synthesize their mutual implications - for my original problem with ecological health as well as for ecological science itself.
The first of these pivotal concepts is Lotka's idea of coupled transformers - two functional types that exist as obligate companion processes, are necessarily interdependent, and are indivisible (Lotka 1925):
"Coupled transformers are presented to us in profuse abundance, wherever one species feeds on another, so that the energy sink of one is the energy source of the other.
A compound transformer of this kind which is of very special interest is that composed of a plant species and an animal species feeding upon the former. The special virtue of this combination is as follows. The animal (catabiotic) species alone could not exist at all, since animals cannot anabolise inorganic food. The plant species alone, on the other hand, would have a very slow working cycle, because the decomposition of dead plant matter, and its reconstitution into CO2, completing the cycle of its transformations, is very slow in the absence of animals, or at any rate very much slower than when the plant is consumed by animals and oxidized in their bodies. Thus the compound transformer (plant and animal) is very much more effective than the plant alone."
Other quotes provide evidence that Lotka worked from a whole-systems perspective as opposed to a focus on mere "interaction" of predator and prey treated as separable parts of a system. For example:
"It is customary to discuss the 'evolution of a species of organisms.' As we proceed we shall see many reasons why we should constantly take in view the evolution, as a whole, of the system (organism plus environment). It may appear at first sight as if this should prove a more complicated problem than the consideration of the evolution of a part only of the system. But it will become apparent, as we proceed, that the physical laws governing evolution in all probability take on a simpler form when referred to the system as a whole than to any portion thereof."
And again later:
"...the concept of evolution, to serve us in its full utility, must be applied, not to an individual species, but to groups of species which evolve in mutual interdependence; and further to the system as a whole, of which such groups form inseparable part."
Thus Lotka suggested evolution as dynamic basis for defining life and ecosystem (Lotka 1925). He also suggested the inseparability of part and whole in living systems lies at the mutually beneficial energy and matter transformations embodied in the plant-animal system taken as a unitary whole. I call his coupled transformers "coupled complementary processes" and see them as a general theme varied multiple times in nature (the main other case being the male-female duality).
In summary, I interpret Lotka's work to argue that 1) coupled transformers are an important dynamical element of living systems, 2) coupled transformers are a general, fundamental theme that is varied many times in nature, and 3) it is best to adopt a holistic treatment of life and its environment as a single evolving system.
Howard Odum depicted the origin of life (Fig. 1) as ecosystem first and cells later (Odum 1970). It is important to note that his origin of life scenario depended on physically-driven cycling within an aquatic system ("circulating seas" as he said). I see Odum's view of the interdependence of production and consumption as Lotka's coupled transformers in another form. Odum's work as a whole has emphasized the primacy of ecosystemic dynamics in understanding life and predicting future development of living systems. His focus on energy as the primary currency of ecosystems reinforces the approach of studying life as interdependent with dynamics of the physical environment.
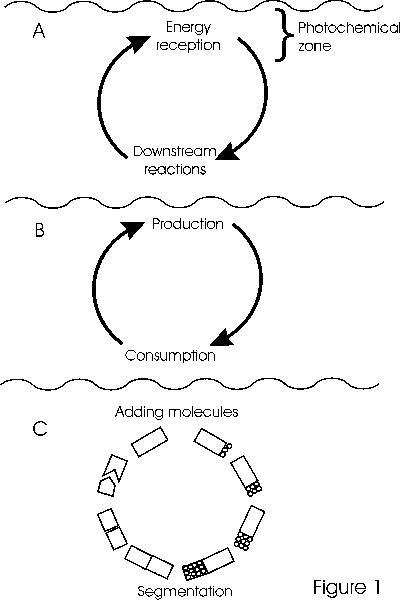
Figure 1. "The ecological system precedes the origin of life", after H.T. Odum (1970).
A. Photoreaction system develops with aid of ocean circulation. B. Storage is developed in polymers as each cycle adds activated molecules. C. Primitive form of reproduction via segmentation of polymers follows. Two additional frames not redrawn show specialization of types of molecular strings and "capsulation and miniaturization" to form cells.
By suggesting a plausible scenario for an ecosystemic origin of life, Odum presents the possibility that ecosystemic functions (e.g., energy flow and nutrient cycling) are more fundamental to life than cellular or organismic functions (e.g., metabolism and reproduction). By extension, he essentially hypothesizes that ecosystemic processes are necessary and sufficient to generate or construct life, cells and organisms from physical and chemical component processes. This constructive aspect qualifies his scenario as an operational definition of life and ecosystem.
In summary, Odum says 1) the ecological system came first and is fundamental, 2) production and consumption were the first differentiation of functions, 3) the origin of life was subsidized by cycling and transport processes of the physical environment.
The third critical concept is ascendency (Ulanowicz 1997). Ascendency is a quantitative measure, derived from information theory, which integrates the magnitude (growth) of system activity as well as the degree of organization (development) among its component processes. Founded on the concept of indirect mutualism, which for Ulanowicz is synonymous with autocatalysis, ascendency captures the dynamics of reaction or trophic networks as they grow, self-organize and complexify. Ulanowicz identifies indirect mutualism as the source of the nonmechanical agency that generates order (through both growth and development) in developing systems. Indirect mutualism is the way in which activity, behavior or variation in performance at one node of a cyclic network eventually returns to the initial node itself and thus either rewards or penalizes that behavior. Through the effects of this causal feedback loop, indirect mutualism imparts key nonmechanical properties to autocatalytic networks, including growth enhancement, selection and competition.
Ulanowicz (1997) also suggests that living systems navigate a "window of vitality" - a middle ground between too much order and too much disorder. He shows this graphically by plotting the effective connectance per node vs topological connectance per node of 38 ecosystems based on their trophic flow networks. All 38 systems exist in a stable region between the extremes of disorder (maximally connected networks, or those at the "edge of chaos") and order (minimally connected, mechanical or "brittle" networks). In its two parts - total system throughput (TST) and average mutual information (AMI) - ascendency models the direction or natural tendency of living system development as a slight asymmetry in a balancing act between ordering and disordering aspects. The TST and AMI terms are multiplied to give ascendency and thus quantify an interaction between generative and constraint. I see the dynamic tension that ascendency captures to be the same as is elsewhere described as the pattern-generating interaction between stimulatory and repressive, activation and inhibition or reaction and diffusion (e.g., Levin and Pacala 1997).
In summary Ulanowicz offers 1) indirect mutualism (autocatalysis) as the key to the systemic, order-generating behavior of life and ecosystems, 2) a dynamic tension between growth and development that gives rise to ascendency, and 3) a "window of vitality" that living systems must navigate between too much and too little order.
Even more than Ulanowicz, Rosen worked with the question "What is life?" directly. But like Ulanowicz, he rethought Newtonian and mechanical paradigms of science in the process. Rosen's (1991) first step in tackling the question was to reframe it. He argued that "Why are organisms different than machines?" is the better question. Thus he built on the assumption that the explanatory powers of the mechanistic worldview cannot help understand the realm of living systems. One of several important clarifications he made was to define a system component as "a unit of organization. A part with a function, i.e., a definite relation between part and whole." From this and other starting concepts, he developed a "relational theory of systems" capable of explaining the special properties of life. Specifically, he identified the "nonfractionability of components in an organism" as the fundamental difference between living systems and machines.
His succinct answer to the big question was: "a material system is an organism if, and only if, it is closed to efficient causation" (Rosen 1991). While this statement and others demonstrate that Rosen considered the fundamental unit of life to be an organism, I think the explanation of this statement and his formal description of the core relation in life still support the ecosystemic life view. In order to produce a model of a system that is closed to efficient cause, Rosen defined replication as a higher order function that can be produced by metabolism and repair functions operating together. Mikulecky (1999) explains how these concepts of Rosen match with the image of life as achieving organizing and disorganizing capacities simultaneously:
"One of the first and most crucial aspects of the evolving living system was its failure to last! It was in a condition of being torn down as fast as it was being built up and this is what allowed it to evolve. Stability is the return to a condition after being perturbed from it. How much more stable could something be than to have both its construction and destruction under strict limits? Both construction and destruction are systems properties. The systematic tearing down allows rebuilding, replication and evolution."
In summary, Rosen argued that 1) life is not divisible like a machine and the mechanistic/reductionist approach will not work to understand it, 2) life is closed to efficient cause, 3) a model of life that meets the requirement of closure to efficient cause includes two integrated functions - metabolism and repair.
I see Rosen's metabolism-repair model as dynamically analogous to Lotka's coupled transformers, Odum's origin via production and consumption and Ulanowicz' growth and development of ecosystems. The ecosystemic life hypothesis exists at the place of congruence of these concepts. Thus the hypothesis is "connectionist" in that it is based upon a network of related concepts. This approach follows Bateson (1979) who sought "the pattern that connects" as a means to understand the unique qualities of living systems. Bateson thought "differences and distinctions" to be key properties of life. Each of the four concepts underlying ecosystemic life includes a dualistic dynamic tension between opposing forces. This shared pattern across fields and between diverse workers inspires the intuitive sense of both life and ecosystem as founded on such dynamic tension at the original and fundamental core.
In the next and final article in this series I will present the ecosystemic life hypothesis itself, explore its implications and discuss its promise.
In the first article in this series (Fiscus 2001), I introduced the ecosystemic life hypothesis as a potential solution to conceptual roadblocks I encountered when trying to define ecological health. In the second article (Fiscus 2002), I presented four concepts that weave life and ecosystem together and suggest that these concepts may be inextricably linked. In this final article, I present the ecosystemic life hypothesis proper and discuss its far-reaching implications.
The ecosystemic life hypothesis melds the ideas of coupled transformers, ecological origin, ascendency, closure to efficient cause and the others mentioned previously into four propositions. I preface the hypothesis using additional concepts of emergence, phase transition and bifurcation but do not define these here; for these I follow the work of Holland (1995), Kauffman (1993) and Prigogine (1996) respectively.
Preface concepts:
i. Life is an emergent property of physical and chemical dynamics. Thus it requires physical and chemical dynamics, but is also independent of those dynamics to a degree described within the concept of emergence.
ii. The original emergence of life from non-living dynamics was a bifurcation into two alternate system states. This bifurcation was also a phase transition in which dynamics began qualitatively new behavior.
The hypothesis:
1. The bifurcation at the origin of life resulted in two functional processes - a molecular string composer (proto-autotroph) and a molecular string decomposer (proto-heterotroph). The composer function is to harness energy (solar, chemical or thermal) to form chemical bonds and build molecular strings. The decomposer function is to harness energy in the chemical bonds and break molecular strings into component parts.
2. The bifurcation into composer and decomposer had the special property that the two system states (or functional subsystems) were interdependent. The composer-decomposer system thus had whole-system capacities that neither subsystem possessed in isolation. These capacities provided a greater ability to survive or persist as a dynamical process in a changing environment - a greater capacity for sustained production.
3. The coupled complementary processes inherent in the composer-decomposer system are necessary and sufficient for life to persist - to sustain physical-chemical dynamical operation - because they counter the two most fundamental threats to life simultaneously. These threats are disorganization (as in dilution) and over-organization (as in crystallization).
4. The ecosystemic organization of life from its origin onward is more fundamental than the cellular or organismic forms of organization. The composer-decomposer system is the common ancestor of both metabolic and genetic processes, both of which are processes of molecular string composition and decomposition. Thus cells were generated later via a process of "encapsulation and miniaturization" (Odum 1970). The ecosystem - autotroph integrated with heterotroph - is the general, self-perpetuating form of life, and cells and organisms are special case subunits of life which cannot persist in isolation.
This hypothesis provides insight into life's capacity for open-ended evolution. The simple functional forms of composer and decomposer operate as a team and in doing so are mutually causal - each helps create and re-create the other (see Macy 1991 on mutual causality). As their operations in concert are iterated many times, varied combinations of molecular building blocks generate the novelty necessary for evolution. The composer-decomposer split is also the genotype-phenotype split, as the composer and decomposer (phenotypes) both also involve molecular strings (genotypes). This is not to say that the molecular strings themselves embody all necessary capacities of life, as I think the cycling first provided by "circulating seas" (Fig. 1, Odum 1970) must have been internalized as well.
Cross-coupling of positive and negative feedbacks in the composer-decomposer system enables life to navigate a window in which entropic (dissipative, disordering) and syntropic (concentrating, ordering; from Fuller 1979) forces are played one against the other and neither dominates. The fact that all known ecosystems depend upon autotrophs and heterotrophs becomes then not an emergent result of interaction or co-evolution, but a unique integration of component part and systemic whole that is the signature basis of life. The hypothesis may be paraphrased thus: life achieves its independence through interdependence. It also suggests that the kernel of the part-to-whole organizational innovation that is the key characteristic of life is the ecosystemic organization of energy flow coupled to nutrient cycling - two topics usually covered in ecology 101.
Because ecosystems have always been conceptualized as fluid in their boundaries with the physical environment, even mutually influential, they provide a better bridge between non-life and life than do the much more clearly bounded units of cell or organism. The ecosystem is thus a plausible intermediate dynamical realm between physics/chemistry and cellular/organismic life. As Solomonoff (1997) suggests, one basis of an operational definition is a sequence of steps for the construction of some entity. Odum provides such a basis for an operational definition of life with his ecological origin scenario. The congruence of his scenario - in the functional coupling between the production and consumption processes - with the concepts of Lotka, Ulanowicz and Rosen enables integration of their whole-systems views of life in such a constructive, operational conceptual framework.
Kauffman (1993) and others have developed the importance of autocatalysis for understanding life and pre-life, but only recently has this concept been extended to highlight the ecosystemic life paradigm. Lee et al. (1997b) talk of "molecular symbiosis" and also "molecular ecosystems" (Lee et al. 1997a) arising from autocatalysis and self-replication.
One way to falsify this hypothesis would be to find a system with only one functional type - either composer or decomposer. As a "thought experiment", the prospect of finding a system with only autotrophs or only heterotrophs seems absurd, but this does not alter the fact that it would refute the claims presented. This gut feeling does show how the idea of ecosystemic life appears at once trivial and profound. Another potential challenge to the hypothesis may be the Archaea - ancient microbes, some of which inhabit hot springs, thermal ocean vents and other such forbidding environments. I admit scant knowledge of the ecological and biological dynamics of this group, and they may well prove the views here wrong.
However, if I were to redo my master's degree research - to start over in my search for an indicator of soil ecological health - I would be much aided by the ecosystemic life hypothesis. In fact, I would approach the problem in a fundamentally different way. First, with help of Ulanowicz' focus on autocatalysis and loops of causality and Lotka's emphasis on whole systems, I would seek initially to sketch a larger, "full-circle" picture of the dynamics in which soils are embedded. As Ulanowicz (1997) shows, the choice of system boundary can make all the difference between seeing dynamics as mechanical versus nonmechanical. From an enlarged view including all major energy and material flows, it becomes clear that nematodes cannot be significant to production or sustainability in conventional agriculture - the subsidies of nutrients provided via fertilizer make their role in mineralization irrelevant. Second, I would seek as benchmark a reference set of dynamics or a reference part-to-whole relationship rather than indices measured at some reference site. In other words, I would not seek to map quantitative measures such as diversity or functional group abundance ratios between reference and study sites, but would compare qualitative, systemic relationships between the two.
Based on this revised approach, I would assess ecological health by the degree to which a system possesses the intact kernel of composer and decomposer subsystems operating in concert. This organizational integrity enables natural systems to operate using renewable energy while also achieving a high degree of materials recycling. All three properties then - composer-decomposer integration, renewable energy and recycling materials processes - are the systemic keys to life's capacities for sustained production and open-ended evolution. This approach thus instigates a shift in perspective away from looking at the nematode communities in soil systems in isolation. Ironically, it is the functional irrelevance of nematodes in the enlarged context of agriculture that helps elucidate the conceptual problems with ecological health. Using a qualitative, systemic criterion for ecological health, the systems with poor health are immediately apparent. Only the human-driven aspects of agricultural systems fail to keep composer and decomposer functions integrated. From this unhealthy systemic organization arise two negative symptoms and a dire prognosis. Conventional agricultural systems decouple composer and decomposer processes and are highly dependent on non-renewable energy and non-recycling materials processes. From the conceptual framework of ecosystemic life, such systems lose the capacity for open-ended evolution and system trajectories are therefore predicted to move toward an evolutionary dead-end. This approach also reveals why I sensed infinite regress in trying to hem in soil ecological health initially. Human disturbance may knock soil systems back in developmental status, complexity, diversity or maturity, but it does not alter the inner source of health. No matter how disturbed by agricultural practices, the life in soils possesses the capacity to restore itself.
Ecosystemic life (as an integrative or "connectionist" concept) may also help resolve recurring conceptual problems within ecology. Three examples of major concepts with which ecosystemic life is consistent simultaneously are: 1) The diversity-stability debate - diversity sometimes helps, sometimes hinders stability; 2) The bottom-up vs top-down ecosystem control debate - ecosystem dynamics are sometimes driven by primary producers, sometimes by higher trophic levels such as keystone species; 3) the unit vs assemblage debate about succession - communities and ecosystems sometimes appear to act like superorganisms, sometimes like assemblies of independent individuals and species. Viewing life as a unified whole requiring two complementary functional units, and thus two inter-twined dynamical aspects, ebbs and flows between diversity and stability, bottom-up and top-down control and unit vs assemblage behavior are to be expected. Ecosystemic life provides a framework by which two seemingly contradictory ecological principles can be true at once, much as the dual or complementarity theory of light does in physics.
I did not cover here all concepts and workers who have contributed to these discussions. I mention Pattee (1973), Maturana and Varela (1980), Root-Bernstein and Dillon (1997), Lovelock (1993) and Margulis et al. (2000) as contributors whose works necessarily must be included in a comprehensive study of these topics.
The ecosystemic life hypothesis inverts the current working assumption that life originated and developed from the cell or organism in the general to the ecosystem in the specific. It also alters the assumption that ecological dynamics are strictly emergent properties arising - from the bottom-up - from interactions of many independent organisms or agents. Instead, organisms are seen as emergent properties of ecological dynamics of energy flow coupled to nutrient cycling. The irreducible or nonfractionable kernel of complexity is the interoperating composer-decomposer system of energy capture and materials cycling. Depew and Weber (1995) state this inversion as the position of a school of thought within systems ecology: "From their perspective, ecosystems are not perspicuously viewed as loosely integrated superorganisms... On the contrary, reformed systems ecologists tend to view organisms as very tightly integrated ecological systems." If this systems ecological approach turns out to be better than an organism-centric approach to understanding life, it could have great implications for how we conduct ecological and life science.
Whole-part integration - the focus on understanding the part-to-whole relationships in living systems - is a synthesis of reductionism and holism. This hybrid paradigm promises new concepts and new approaches to old problems that can enable meaningful development of the concept of ecological health, as well as other benefits perhaps imaginable. For example, could the ecosystemic life and part-to-whole approach enable design of human systems such that emergent properties, while not fully controllable or predictable, can be steered toward the beneficial? The ultimate benchmark for system health and quality, as well as the ultimate role model, mentor and design guide for human policy and development realms, may well be life itself.
Thanks to Bob Ulanowicz for valuable comments on an earlier version of this three-part essay. Thanks also to Pat Kangas for helpful discussion and for the reference to H.T. Odum (1970).
Bateson, G. 1979. Mind and Nature: A Necessary Unity. Bantam Books, New York, NY. USA.
Britannica online encyclopedia at http://www.britannica.com/.
Depew, D.J. and B. H. Weber. 1995. Darwinism Evolving: Systems Dynamics and the Genealogy of Natural Selection. MIT Press, Cambridge, MA. USA.
Eigen, M. 1995. What will endure of 20th century biology? In M.P. Murphy and L.A.J. O'Neill, Eds. What is Life? The Next Fifty Years: Speculations on the future of Biology. Cambridge University Press. Cambridge. England.
Ettema, C.H., and T. Bongers. 1993. Characterization of nematode colonization and succession in disturbed soil using the Maturity Index. Biology and Fertility of Soils 16:79-85.
Fiscus, D.A. 2001. The ecosystemic life hypothesis I: introduction and definitions. Bulletin of the Ecological Society of America 82(4): 248-250, October 2001.
Fiscus, D.A. 2002. The ecosystemic life hypothesis II: four connected concepts. Bulletin of the Ecological Society of America, January, 2002.
Fiscus, D.A. and D.A. Neher. In press. Distinguishing sensitivity of free-living soil nematode genera to physical and chemical disturbances. To appear in Ecological Applications.
Fuller, R.B. 1979. Synergetics 2: Explorations in the Geometry of Thinking. MacMillan Publishing Co., New York, NY. USA.
Hess, G. R., C. L. Campbell, D. A. Fiscus, A. S. Hellkamp, B. M. McQuaid, M. L. Munster, S. L. Peck, and S. R. Shafer. 2000. A conceptual model and indicators for assessing the ecological condition of agricultural lands. Journal of Environmental Quality 29:728-737
Holland, J. 1995. Hidden Order: How Adaptation Builds Complexity. Addison-Wesley, Reading, MA. USA.
Karr, J.R. 1991. Biological integrity: a long-neglected aspect of water resource management. Ecological Applications 1:66-84.
Kauffman, S.A. 1993. The Origins of Order. Oxford University Press. New York, NY. USA.
Lee, D.H., K. Severin and M.R. Ghadiri. 1997a. Autocatalytic networks: the transition from molecular self-replication to molecular eosystems. Current Opinion in Chemical Biology 491-496.
Lee, D.H., K. Severin, Y. Yokobayashi, and M.R. Ghadiri. 1997b. Emergence of symbiosis in peptide self-replication through a hypercyclic network. Nature 390:591-594.
Levin, S.A. and S.W. Pacala. 1997. Theories of simplification and scaling of spatially distributed processes. In D. Tilman and P. Kareiva Eds. Spatial Ecology. Princeton University Press. Princeton, NJ. USA.
Lotka, A.J. 1925. Elements of Physical Biology. Wilkins and Wilkins, Baltimore, MD. USA.
Lovelock, J.E. 1993. The soil as a model for the earth. Geoderma 57(3): 213-215.
Macy, J. 1991. Mutual Causality in Buddhism and General Systems Theory: The Dharma of Natural Systems. SUNY Press, Albany, NY. USA.
Margulis, L., C. Matthews and A. Haselton. 2000. Eds. Environmental Evolution: Effects of the Origin and Evolution of Life on Planet Earth. MIT Press, Cambridge, MA. USA.
Maturana, H.R. and F.J. Varela. 1980. Autopoesis and Cognition: The Realization of the Living. D. Reidel Publishing Co., Dordrecht, Holland.
Merriam-Webster Online dictionary at http://www.m-w.com/dictionary.htm.
Mikulecky, D.C. 1999. Robert Rosen: The well posed question and its answer - why are organisms different from machines? Online article available at http://views.vcu.edu/~mikuleck/
Morowitz, H.J. 1992. The Beginnings of Cellular Life: Metabolism Recapitulates Biogenesis. Yale University Press. New Haven, CT. USA.
Odum, H.T. 1970. Environment, Power and Society. Wiley-Interscience. New York, NY. USA.
O'Neill, R.V., D.L. DeAngelis, J.B. Waide, T.F.H. Allen. 1987. A Hierarchical Concept of Ecosystems. Princeton, NJ: Princeton University Press. 253 pp.
Pattee, H.H. 1973. Ed. Hierarchy Theory: The Challenge of Complex Systems. George Braziller, New York, NY. USA.
Prigogine, I. 1996. End of Certainty: Time, Chaos and the New Laws of Nature. The Free Press. New York, NY. USA.
Root-Bernstein, R.S. and P.F. Dillon. 1997. Molecular complementarity I: the complementarity theory of the origin and evolution of life. Journal of Theoretical Biology 188(4):447-479.
Rosen, R. 1991. Life Itself. Columbia University Press. New York, NY.
Schrodinger, E. 1944. What is Life? The Physical Aspect of the Living Cell. Cambridge University Press, Cambridge, MA. USA.
Solomonoff, R.J. 1997. The discovery of algorithmic probability. Journal of Computer and System Sciences 55(1):73-88.
Ulanowicz. R.E. 1997. Ecology, the Ascendant Perspective. Columbia University Press. New York, NY. USA.